what two conditions does the closing of the stomata help to prevent
How do plants breathe through stomata? Key regulators of stomata are plant vacuoles, fluid-filled organelles leap by a single membrane called the tonoplast.
Similar animals, plants breathe. The gas exchange into and out of a plant leaf occurs at the underside of leaves, and the process is precisely regulated. What are the gases that are exchanged at the leaf surface? The main free energy-producing biochemical process in plants is photosynthesis, a process that, initiated by energy from the sun, converts CO2 and water into carbohydrate energy molecules for the plant and releases O2 back into the atmosphere. In this process, leaves take in atmospheric COtwo and release O2 back into the air. How practise plants perform these gas exchange activities betwixt leaf cells and the outside surroundings? Scientists discovered that a distinct organelle, the vacuole, plays a critical office in regulating the commitment of COii to the photosynthesis-conducting chloroplasts.
Plant vacuoles are fluid-filled organelles leap past a single membrane called the tonoplast, and contain a wide range of inorganic ions and molecules. Scientists accept identified at least two types of plant vacuoles. The two primary types are the poly peptide storage vacuoles of neutral pH, and the lytic vacuoles of acidic pH, which are equivalent in function to lysosomes in mammalian cells (Figure i).
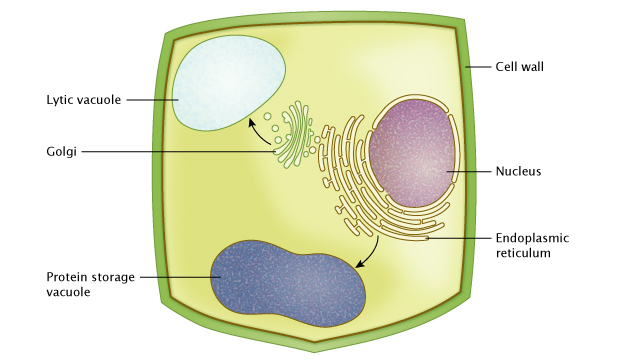
Figure 1: Vacuoles in plant cells
Vacuolar proteins are synthesized and processed in the endoplasmic reticulum (ER), and transferred to vacuoles through diverse routes. They can transfer indirectly via the Golgi appliance to a lytic vacuole. They tin also transfer directly from the endoplasmic reticulum (ER) to a poly peptide storage vacuole. As a prison cell grows, protein vacuoles can gradually fuse with each other and form much larger vacuoles (not shown).
Vacuole Size Changes Are Correlated with Stomatal Movements
During photosynthesis, leaves accept in atmospheric COii and release O2 through stomata, microscopic pore structures in the leaf epidermis (atypical = stoma). A pair of guard cells surrounds each stoma, and these cells command the opening and closing of the stomatal pore between them. Guard cells regulate this opening and closing in response to a wide multifariousness of environmental signals, such as day/nighttime rhythms, CO2 availability, and temperature. Why do plants spend energy on opening and closing these stomata, when they could leave them constantly open up, and let CO2 flow freely? The primary reason is that stomata also regulate the passage of h2o molecules. If the stomata were constantly open, plants would lose too much h2o via evaporation from the leaf surface, a process chosen transpiration.

Figure 2: The expression of green fluorescent protein (GFP) fused with a tonoplast protein AtCLCa, showing the boundary of tonoplast in protoplasts.
Left, laser-scanning confocal microscopy prototype of GFP fluorescence; Center, transmitted lite microscopy image; Right, merged prototype of both; calibration bars correspond to xvi µm.
© 2013 Nature Publishing Grouping De Angeli, A. et al. The nitrate/proton antiporter AtCLCa mediates nitrate accumulation in plant vacuoles. Nature 442, 939–942 (2006) doi:10.1038/nature05013. All rights reserved. ![]()
A special characteristic of baby-sit cells is that they tin can increase or decrease their book, thereby irresolute their shape. This is the basis for the opening and closing of a stoma, known as stomatal movement, which controls gas exchange necessary for photosynthesis and limits water loss. How practice guard cells change their volume to control this opening and endmost? They exercise so by irresolute the osmotic force per unit area of their vacuoles, which then either have upwardly or lose water, and consequently enlarge or shrink. Such changes in vacuolar book are quite rapid and dramatic. This can exist problematic because, unlike a quickly expanding balloon, biological membranes are more limited in their elasticity and practice not allow over-stretching. How and then does the tonoplast increase its area so that the vacuole can quickly expand (Figure two)? Scientists still know very piddling near this dynamic process, and they are actively searching for a mechanism, 1 that peradventure provides reservoirs of membrane to the tonoplast to adjust such a rapid volume change.
How Do Vacuoles Change During Stomatal Opening and Closing?
One way to rails dynamic changes in guard cell vacuoles during stomatal movements is to utilize cell imaging techniques, such as confocal microscopy and TEM. In 2005, Gao et al. did just this when they studied leafage epidermis from the plant Vicia faba using microscopy coupled with fluorescent dyes. Offset, they removed strips of epidermal cells from leaves, so they stained guard cells with various fluorescent dyes. They used two dyes that specifically attach to vacuoles due to their acidic pH. These dyes cause the vacuoles to glow fluorescent dark-green or red. They also used a green dye that remains in the cytoplasm and does not enter vacuoles. This dye gives an inverse image to the vacuole-specific dyes (Figure iii). With the use of these compartment-specific dyes, they were able to observe the size, shape, and number of vacuoles at various time points during stomatal movements. In their experiment, Gao et al. asked, what happens to the vacuoles and the cytoplasm during stomatal opening and endmost? They controlled stomatal action experimentally with known agents. They induced opening with halogen cold-light, and closing with chemical abscisic acid (ABA). During these inductions, they observed that, in the closed state, guard cells contain many minor vacuoles, only during stomatal opening, these pocket-sized vacuoles readily fuse with each other, or with bigger vacuoles. The outcome is very big vacuoles in guard cells surrounding an open stoma. Conversely, in endmost stomata, the large vacuoles once again split into smaller ones, and generate many circuitous membrane structures. Though these scientists observed a visual coincidence of vacuole changes and stomatal movements, are these dynamic changes necessary for stomatal movements to occur?
Does Stomatal Opening Crave Vacuolar Fusion?
To test whether vacuole dynamics are necessary, Gao et al. asked, what would happen to stomatal movements if they experimentally disrupt vacuolar fusion? To investigate this problem, they again turned to their test organisation, leaf epidermal peels. They treated these peels with a membrane-permeable chemical compound known to inhibit the fusion of endosomes with vacuoles, called E-64d ((2s,3s)-trans-epoxy-succinyl-50-leucylamido-3-methylbutane ethyl ester), and establish that the treated guard cells had a greater number of vacuoles than untreated command guard cells. They besides observed that stomatal opening was slower in treated guard cells compared to the untreated controls. To explain this, they concluded that interrupted vacuolar fusion has an effect of slowing stomatal opening, and therefore vacuolar fusion must exist necessary for stomatal opening to properly office. To explore the genetic basis for vacuolar dynamics, Gao et al. followed up this initial conclusion in Vicia faba with additional experiments using mutant plants. Genetic manipulation in the establish Arabidopsis can produce a mutant that is defective in producing a poly peptide named SGR3. Previous work by other scientists established that SGR3 impacts the transport of vesicles into vacuoles and vacuolar fusion. When Gao et al. compared SGR3 mutants to normal (wild type) plants, they found slower stomatal opening in response to light induction in the mutant plants. With their knowledge of SGR3 function, and these observations, they again ended that impaired stomatal motion was a consequence of reduced vacuolar fusion in guard cells. Altogether, their results show that fusion of vacuoles is necessary for normal, rapid stomatal movements.
What Are Other Mechanisms for Vacuolar Expansion?
Are there other ways guard cells can increment vacuolar volume bated from fusing small vacuoles? The answer appears to exist yep. Until recently, researchers believed that the tonoplast (vacuolar membrane) had a smooth surface. Modern jail cell imaging techniques with live plant cells have shown otherwise. With confocal microscopy of live cells, several research groups have observed a wavy vacuolar surface, called tonoplast foldings, and vesicle-similar structures within the vacuolar lumen (Cutler et al. 2000; Verbelen & Tao 1998; Yamamoto et al. 2003). Using time-lapse imaging, Gao et al. also observed these foldings and vesicle-like structures, which disappeared upon stomatal opening merely re-appeared during stomatal closing. This led them to conclude that these observed intravacuolar membrane structures may serve as reservoirs for the tonoplast, so it has membrane set up to supply any needs for rapid expansion. Furthermore, they discovered that what at first appeared to exist private vacuoles might actually exist connected ones. They discovered this when they directed a strong axle of excitation light directly onto a single vacuole containing fluorescent dye. This precise axle of light causes photobleaching of dye molecules, and they afterward observed photobleached dye molecules in both the targeted vacuoles and neighboring ones, indicating that in that location may be some physical continuity between what appeared to be dissever vacuoles. The scientists ended that neighboring vacuoles are physically linked. Altogether, these experimental observations propose that vacuoles, tonoplasts, and intravacuolar membrane systems interact to sustain rapid stomatal movements, a prerequisite for CO2 uptake during photosynthesis.
Many questions remain about the exact mechanisms of vacuole expansion that regulate stomatal movements. For instance, why does guard cell volume increase involve expansion of a few big vacuoles instead of the commonage expansion of many smaller ones? Fusion of large vacuoles tin swallow a large amount of energy than the expansion of multiple smaller ones. A possible caption for this seemingly inefficient energy strategy lies in the tradeoff for volume over surface area. A big vacuole would have a greater total volume/surface area ratio than many pocket-sized ones. This makes a large vacuole more effective at volume expansion.
Another question is, what is the exact mechanism of vacuolar fusion, and nether what conditions does information technology occur? Many environmental factors affect stomatal motility, and one of them is CO2, the very gas that enters stomata. With the ever-accumulating reports of increasing COii concentration in our atmosphere, these questions almost stomatal regulation have relevance to issues of global climate change. How volition this increasing atmospheric CO2 concentration influence the regulation of baby-sit prison cell movement? Scientists are currently investigating these phenomena with sophisticated genetic tools and microscopy.
Summary
Plant vacuoles are ubiquitous organelles that are essential to multiple aspects of plant growth, maintenance and development. Their key part in stomatal movements underscores their importance in fundamental gas exchange for plants. Scientists are actively pursuing the exact mechanisms that control vacuole fusion, which supports stomatal movements too as other constitute cell functions. In improver to their role in controlling photosynthetic gas exchange, vacuoles likewise store compounds that may help to protect photosystems in the chloroplast from damage acquired by backlog light. Vacuoles are important compartments in plant cell metabolism. An intact vacuole is necessary for many plant functions. Scientists are working toward identifying and characterizing a large and various group of tonoplast transporters. They ask: What is their molecular structure? What do they do? How might they exist linked? Understanding the answers to these questions is important, equally they show how vacuoles are an integrated component of complex cellular networks. Every bit the data in a higher place show, vacuoles are crucial to establish cells, as they enable gas substitution mechanisms that optimize metabolic atmospheric condition in the cytosol, and let a plant to react to changing ecology conditions.
References and Recommended Reading
Allen, G.J. et al. A divers range of guard cell calcium oscillation parameters encodes stomatal movements. Nature 411, 1053–1057 (2001)
Brandizzi, F. & Hawes, C. A long and winding road. EMBO reports 5, 245-249 (2004). doi: 10.1038/sj.embor.7400099
Cutler, South.R. et al. Random GFP::cDNA fusions enable visualization of subcellular structures in cells of Arabidopsis at a high frequency. Proc Natl Acad Sci USA 97, 3718–3723 (2000)
Gao, X. et al. The Dynamic Changes of Tonoplasts in Guard Cells Are Important for Stomatal Movement in Vicia faba. Plant Physiology 139, 1207–1216 (2005)
Gilroy, S. & Trewavas, A. Signal processing and transduction in plant cells: the end of the showtime? Nature Reviews Molecular Prison cell Biological science ii, 307–314 (2001) doi:10.1038/35067109.
Hu, H. et al. Carbonic anhydrases are upstream regulators of COii-controlled stomatal movements in baby-sit cells. Nature Jail cell Biology 12, 87–93 (2009) doi:x.1038/ncb2009.
Lee, M. et al. The ABC transporter AtABCB14 is a malate importer and modulates stomatal response to CO2. Nature Prison cell Biology 10, 1217–1223 (2008) doi:ten.1038/ncb1782.
Peters, C. & Mayer, A. Ca2+/calmodulin signals the completion of docking and triggers a belatedly step of vacuole fusion. Nature 396, 575–580 (1998)
Sanderfoot, A. A. et al. The t-SNARE AtVAM3p resides on the prevacuolar compartment in Arabidopsis root cells. Plant Physiol 121, 929–938 (1999)
Sato, M et al. The AtVAM3 encodes a syntaxin-related molecule implicated in the vacuolar assembly in Arabidopsis thaliana. J Biol Chem 272, 24530–24535 (1997)
Serna, L. Coming closer to a stoma ion channel. Nature Cell Biology 10, 509–511 (2008) doi:10.1038/ncb0508–509.
Schroeder, J. I. et al. Guard cell abscisic acid signalling and applied science drought hardiness in plants. Nature 410, 327–330 (2001)
Surpin, Chiliad. & Raikhel, North.. Traffic jams impact plant development and signal transduction. Nature Reviews Molecular Cell Biological science v, 100–109 (2004) doi:10.1038/nrm1311.
Verbelen, J.P. & Tao, W.. Mobile arrays of vacuole ripples are common in plant cells. Found Prison cell Rep 17, 917–920 (1998)
Yamamoto, Y. et al. Beliefs of vacuoles during microspore and pollen development in Arabidopsis thaliana. Establish Jail cell Physiol 44, 1192–1201 (2003)
Source: https://www.nature.com/scitable/topicpage/plant-vacuoles-and-the-regulation-of-stomatal-14163334/
0 Response to "what two conditions does the closing of the stomata help to prevent"
Post a Comment